
First published in Nature Vol. 207, No. 4997, pp. 568-570, August 7, 1965.
The design of an efficient and unequivocal experiment [1] in extra-terrestrial life detection should take into account :
- A definition of life stated in terms favourable for its recognition.
- A description of tho past and present environment of the planet to be sampled.
As yet, there is no formal physical statement to describe life from which an exclusive definition for experimental purposes could be drawn. Moreover no comprehensive description is available of the atmospheric as well as the surface physical and chemical environment of any of the planetary bodies.
It is not surprising, in view of the vast expense of space-probe experiments and of the formidable uncertainties already stated here, that the proposed experiments in life detection all ask the cautious geocentric question: “Is there life as we know it ?” Most certainly it is difficult to envisage in detail an alien biochemistry; it would seem pointless and very uneconomic to send a space probe to detect a speculative life-form.
It is the object of this article to show that we are not necessarily limited to experiments based on the recognition of a specific life-form, either Earth-like or alien. Also, that it is possible, by accepting a limited phenomenological definition of life, to design simple experiments from the general recognition of life phenomena, including that with which we are familiar. The application of this approach to experiments in life detection is the basis of the discussion which follows.
Recognition of Life
It is a relatively simple matter to distinguish between living and inorganic matter on Earth by biochemical experiments even though no formal definition of life in biochemical terms exists. Experience suggests, for example, that a system capable of converting water, atmospheric nitrogen and carbon dioxide into protein, using light as a source of energy, is unlikely to be inorganic. This approach for recognition of life by phenomenology is the basis of the experiments in detection of life so far proposed. Its weakness lies not in the lack of a formal definition but in the assumption that all life has a common biochemical ancestry.
It is also possible to distinguish living from inorganic matter by physical experiments. For example, an examination of the motion of a salmon swimming upstream suggests a degree of purpose inconsistent with a random inorganic process. The physical approach to recognition of life is no more rigorous, at this stage, than is the biochemical one; it is, however, universal in application and not subject to the local constraints which may have set the biochemical pattern of life on Earth.
Past discussions of the physical basis of life [1 – 3] reach an agreed classification as follows:
‘‘Life is one member of the class of phenomena which are open or continuous reaction systems able to decrease their entropy at the expense of substances or energy taken in from the environment and subsequently rejected in a degraded form.’’
This classification is broad and includes also phenomena such as flames, vortex motion and many others. Life differs from the other phenomena so classified in its singularity, persistence, and in the size of the entropy decrease associated with it. Vortices appear spontaneously but soon vanish; the entropy decrease associated with the formation of a vortex is small compared with energy flux. Life does not easily form, but ‘persists indefinitely and vastly modifies its environment. The spontaneous generation of life, according to recent calculations from quantum mechanics [4, 5], is extremely improbable. This is relevant to the present discussion through the implication that wherever life exists its biochemical form will be strongly determined by the initiating event. This in turn could vary with the planetary environment at the time of initiation.
On the basis of the physical phenomenology already mentioned, a planet bearing life is distinguishable from a sterile one as follows:
- The omnipresence of intense orderliness and of structures and of events utterly improbable on a basis of thermodynamic equilibrium.
- Extreme departures from an inorganic steady-state equilibrium of chemical potential.
This orderliness and chemical disequilibrium would to a diminished but still recognizable extent be expected to penetrate into the planetary surface and its past history as fossils and as rocks of biological origin.
Experiments for Detection of Life
The distinguishing features of a life-bearing planet, described here, suggest the following simple experiments in detection of life:
A. Search for order.
- Order in chemical structures and sequences of structure. A simple gas chromatograph or a combined gas chromatograph – mass spectrometer instrument would seek ordered molecular sequences as well as chemical identities.
- Order in molecular weight distributions. Polymers of biological origin have sharply defined molecular weights, polymers of inorganic origin do not. A simple apparatus to seek ordered molecular weight distributions in soil has not yet been proposed but seems worthy of consideration.
- Looking and listening for order. A simple microphone is already proposed for other (meteorological) purposes on future planetary probes; this could also listen for ordered sequences of sound the presence of which would be strongly indicative of life. At the present stage of technical development a visual search is probably too complex ; it is nevertheless the most rapid and effective method of life recognition in terms of orderliness outside the bounds of random assembly.
B. Search for non-equilibrium.
- Chemical disequilibrium sought by a differential thermal analysis (DTA) apparatus. Two equal samples of the planetary surface would be heated in a DTA apparatus: one sample in the atmosphere of the planet, the other in an inert gas, such as argon. An exotherm on the differential signal between the two samples would indicate a reaction between the surface and its atmosphere, a condition most unlikely to be encountered{ where there is chemical equilibrium as in the absence of life. It should be noted that this method would recognize reoxidizing life on a planet with a reducing atmosphere. This experiment could with advantage and economy be combined with, for example, the gas chromatography mass spectrometry experiment (Al) where it is necessary to heat the sample for vaporization and pyrolysis.
- Atmospheric analysis. Search for the presence of compounds in the planet’s atmosphere which are incompatible on a long-term basis. For example, oxygen and hydrocarbons co-exist in the Earth’s atmosphere.
- Physical non-equilibrium. A simplified visual search apparatus programmed to recognize objects in non-random motion. A more complex assembly could recognize objects in metastable equilibrium with the gravitational field of the planet. Much of the plant life on Earth falls into this category.
Experiments A1, B1 and B2 are the most promising for the development of practical instruments. Indeed, the gas chromatography – mass spectrometry combination experiment and the DTA experiment already proposed for planetary probes [7] are, with minor modifications, capable of recognizing the ordered sequences and chemical disequilibrium discussed earlier. Experiment B2, atmospheric analysis, is simple and practical as well as important in the general problem of detection of life. A detailed and accurate knowledge of the composition of the planetary atmosphere can directly indicate the presence of life in terms of chemical disequilibrium; such knowledge also is complementary to the understanding of other life.
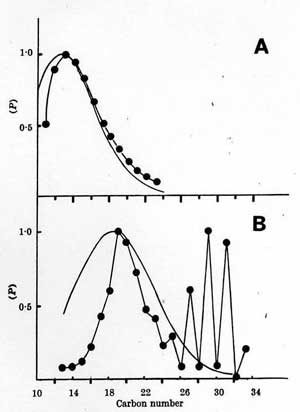
Fig. 1. Caption: The abundance of n-alkanes from an inorganic source (A), Fischer-Tropsch hydrocarbons, and from a biological source (B), wool wax. The observed abundances (o-o) are compared with normalized Poisson distributions (-) around the preponderant alkanea detection experiments and to the planning of subsequent experiments. Even on Earth whore life is abundant there are many regions, such as those covered by fresh snow, where a surface sample might be unrewarding in the search for life. The atmospheric composition is largely independent of the site of sampling and provides an averaged value representative of the steady state of chemical potential for the whole planetary surface.
Fig. 1 shows the abundance of hydrocarbons of carbon number between 11 and 33 for abiotic hydrocarbons of the Fischer-Tropsch process [8] and hydrocarbons of biological origin, wool wax [9]. Poisson distributions around the predominant hydrocarbon numbers are shown as solid lines. The inorganic hydrocarbons fit closely the expected Poisson distribution for a state of chemical equilibrium. By contrast the biological hydrocarbons show large departures in the distribution of their abundance from this equilibrium state; also, especially for the higher molecular weight alkanes, a two-carbon ordered sequence is well established.
In a similar manner with experiment B1 the disequilibrium associated with life can be demonstrated. A few mg of soil heated in a DTA apparatus in air shows a large exotherm when compared with a similar reference sample heated in argon. The combustion of even a few micrograms of organic matter in these circumstances is capable of generating a detectable signal.
Detection of Life on Mars
Ordinarily one does not look for fish in a desert, nor for cacti on. an ice cap. Should we, therefore, look for microorganisms of Earth-like habits on Mars, or should we rather ask one or more of the general questions discussed here? The answer to this must depend on the history of Mars, past and present.
The following is the only sure information so far available on Mars. It is dry. The atmosphere is thin and contains no more than a trace of oxygen. The flux of solar radiation at the surface, although less than on Earth, is also less filtered and may include an appreciable content of energetic radiation; in particular, short wave-length ultra-violet. The temperature range includes periods above zero centigrade. Finally, but less certain, is the possibility [6] that oxides of nitrogen are present in appreciable amounts.
If these conditions are representative of Mars in the past as well as now, there seems no reason to assume that life, if present at all, can resemble that on Earth. However, it is possible that Mars was once Earth-like (primeval non-living Earth) and has changed physically to its present state; or less likely that the present state of Mars, like that of the Earth, is a consequence of biological change. For success a geocentric biochemical experiment must assume that Mars was once Earth-like and that life is still surviving in a highly adapted form yet still recognizable to the experiment.
If Mars is as it always was or has been changed to its present state by biological action, then life, if there now, would be very different from that we know. Could we conceive of living systems in liquid N2O4, as an ionizing solvent ? Could they use hard ultra-violet as a source of energy ? Is cellular life necessary in a dry environment, or did cell membranes evolve on Earth to offset the overwhelming effects of dilution in the primeval seas ? What sort of Martian biochemistry could have generated the present atmosphere ?
Answers to these questions are important in the design of experiments to detect particular life-forms; thus, with a growth experiment, or a biochemical experiment, the strength, composition and conditions of incubation of the medium are of vital importance. This information, however, is not needed in the general detection of life. In view of what is not known of conditions on Mars, the physicochemical experiments in life recognition such as experiments A1 and B1 and B2 seem more worth considering for early probe experiments. These simple experiments do not require a prior knowledge of the planetary environment and are not limited to Earth biochemistry.
Acknowledgements
I thank A. Zlatkis and P. G. Simmonds of the University of Houston for their advice and for conducting and providing me with the results of the differential thermal analysis experiments suggested in the discussions here. I also thank G. Hobby and G. Mamikunian of the Jet Propulsion Laboratory, California Institute of Technology, Pasadena, California, for their advice.
This work was supported by a grant from the National Aeronautics and Space Administration (NSG 199-62, J. E. Lovelock).
References
1. Schrodinger, E., What is Life (Camb. Univ. Press, 1944).
2. Bernal, J. D., The Physical Basis of Life (Routledge and Kegan Paul, London, 1951).
3. Denbigh, K. G., The Thermodynamics of the Steady State (Methuen and Co., London, 1951).
4. Wigner, E. P., The Logic of Personal Knowledge (Routledge and Kegan Paul, London, 1961).
5. Landsberg, P. T., Nature, 203, 928 (1964).
6. Kiess, C. C., Karrer, S., and Kiess, K., Publ. Astro. Soc. Pacific, 72, 256 (1960).
7. Lipsky, S. R., and Lovelock, J. E., Recommendations report. Submitted to Donald Easter, Planetary Atmospheres Section, NASA Headquarters, Washington, D.C. (April 1964).
8. Meinschein, W. G., Space Sci. Revs., 2, 665 (1963).
9. Mold, J. D., et al., Biochemistry, 3, 1293 (1964).