
By J. E. Lovelock, Coombe Mill Experimental Station, St Giles on the Heath, Launceston, Cornwall, England PL15 9RY.Published in P. Westbroek and E. W. deJong (eds.), Biomineralization and Biological Metal Accumulation, pp.15-25. Copyright © 1983 by D. Reidel Publishing Company.
Abstract. Life can flourish only within a narrowly circumscribed range of physical and chemical states and since life began the Earth has kept within this range. This is remarkable for there have been major perturbations such as a progressive increase in solar luminosity, extensive changes in the surface and atmospheric chemical composition and the impact of many planetesimals. The anomalous and chemically unstable composition of the Earth’s atmosphere when compared with those of the other terrestrial planets was the first indication of homeostasis by the biota to maintain conditions favourable for their continued survival. This paper will discuss recent evidence in support of the Gaia hypothesis and present a simple model of a planetary ecosystem in which homeostasis is a direct and automatic result of the characteristic properties of life.
It is now just over ten years since the first paper on the Gaia hypothesis was published. Lovelock and Margulis (9) . Many investigators including Redfield (13), Hutchinson (8) and Sillen (15) had previously noted the strong influence of the biota on the composition and properties of the Earth. Like these earlier attempts to unify the biological and geochemical approaches to understanding the Earth, the Gaia hypothesis has tended to be ignored rather than criticised by geochemists, almost as if Aristotle still ruled and anything moving towards a circular, even a nonlinear, argument was forbidden. Gaia, which uses the circular reasoning of cybernetics, was taken to be teleological. Two biologists, Ford Doolittle (4) and Dawkins (3) were prepared to criticise the hypothesis openly on the grounds that it was contrary to the expectations of natural selection but even they did not or were unable to express their views in main stream scientific literature.
The purpose of this paper is to try to demystify the Gaia hypothesis and to illustrate it by a simple model which is in a form more acceptable for discussion and criticism. The discussion will be drawn mostly from obervations about the composition, properties and regulation of the atmosphere. Lynn Margulis provides in the next paper a parallel account of the Gaia hypothesis taken in the context of the biogeochemistry of the sediments.
What is Gaia?
The Gaia hypothesis arose directly from the planetary exploration programme of NASA. There was a need to discover in advance of a landing mission whether or not a planet such as Mars bore life. In 1966 Hitchcock and Lovelock (6) were able to show that information on the atmospheric composition of a planet was sufficient as prima facia evidence of life. The method was based on the high probability that planetary life, through its use of the atmosphere, would drive the chemical composition of this medium far from the near equilibrium steady state of a lifeless planet. This detection method when applied to Mars strongly indicated it to be barren, a conclusion highly unacceptable to exobiologists at that time. The same method applied to the Earth indicated the near certainty of the presence of life. It also suggested that the atmosphere was more than just a biogeochemical mixture. It appeared to be actively maintained in composition at close to an optimum by and for the biota. This way of thinking about the planets was a stunning discouragement for exobiologists whose scientific inspiration came from the search for life, outside the Earth. Some part of the tendency to ignore the joyous counterpart of this approach, the discovery of Gaia perhaps arose from their disappointment.
The evidence drawn from atmospheric compositions which points to life and to a control system on a planet is summarised in Figures 1 and 2, which show the abundances and the fluxes respectively, of the gases of the present atmosphere compared with those of an abiological Earth. From these diagrams it is clear as has been argued in previous papers (9,10,12) that the atmosphere is a highly reactive mixture which would but for life rapidly revert to the stable inert condition of the abiological state. It is the intense disequilibrium of the atmosphere which advertises the presence of life on Earth. It is the maintenance of this reactive and unstable atmosphere at a steady state for times much longer than the residence times of individual gases that suggests the presence of a control system, Gaia.
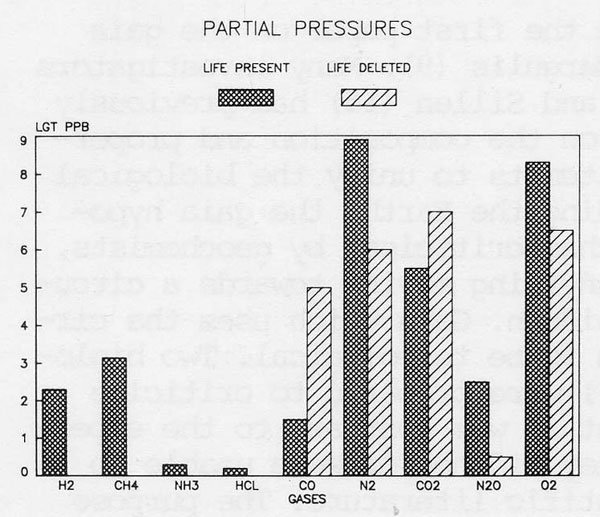
Figure 1. The abundance of gases in the present atmosphereic gas flux compared with that expected of the abiological steady state.
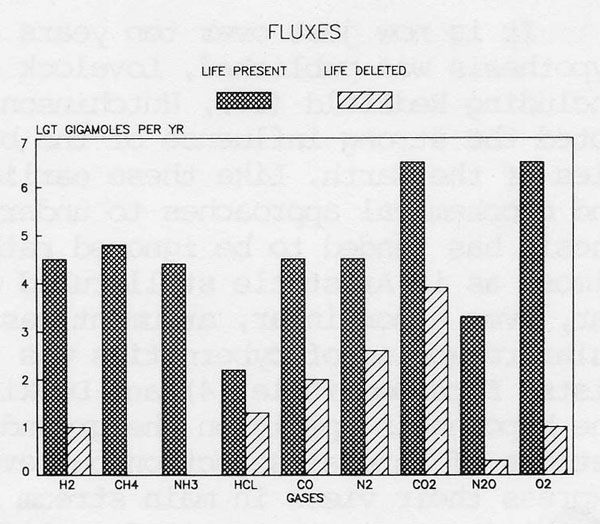
Figure 2. The fluxes of gases (gigamoles per year) through the present atmosphere compared with those expected for the abiological steady state.
Feedback and homeostasis
Many geochemists now accept that the Earth’s surface features are a result of the coevolution of the biota and the rocks. But they still see the association between life and its environment as passive. Life adapts to environmental change and the evolution of life may change the environment but any feedback, negative or positive between these processes is passive. In sharp contrast the Gaia hypothesis sees the Earth as homeostatic, with the biota actively seeking to keep the environment optimal for life. Such a view is usually regarded as teleological or mechanistic, Holland (7), and Gilbert (5).
The first intimation of Gaia comes from the realisation that every evolutionary step of the biota must to a greater or lesser extent alter the environment in which the next generation will evolve. If the change is to a more favourable environment then it will carry more progeny and the environmental change will be reinforced. In the same way a less favourable environment will carry less progeny and hence their unfavourable attributes will become attenuated.
It is not immediately obvious how such a course of events could lead to planetary homeostasis. As Ford Doolittle (4) observed the biota have no capacity for conscious foresight or planning and would not in the pursuit of local selfish interests evolve an altruistic system for planetary improvement and regulation.
The sequential logic of descriptive writing is not designed for the concise explanation of control systems with their inherent circularity recursiveness and non-linearity. Even the formalism of mathematics loses its elegance when an attempt is made to describe a simple nonlinear control system such as, for example, an electrical water heater controlled by a bimetallic strip thermostat. I have chosen therefore to present a simple model of an imaginary planet whose temperature is regulated at a biological optimum over a wide range of solar radiation levels as a working example of a Gaian mechanism.
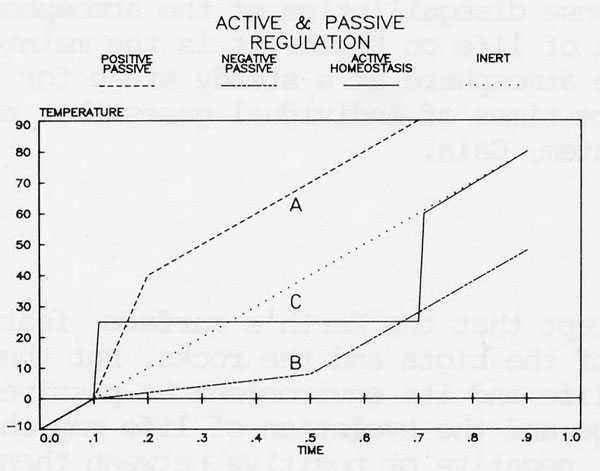
Figure 3. Diagram to illustrate passive negative and positive feedback processes and to compare them with active hcmeostasis. Energy is supplied at a constant rate to an object of unknown thermal mass. The diagonal line across the diagram illustrates the rate of temperature rise of an inert object. Line (B) illustrates passive negative feedback on the energy supply over a limited range. Line (A) passive positive feedback over the same range and line (C) active regulation at a chosen set temperature.
Before describing this model it is useful first to consider the terms active, passive and feedback in the context of their origin, namely systems engineering. Figure 3 illustrates graphically the change of some intrinsic property of a system, such as temperature, with time when there is a constant flux of a related quantity such as heat. The diagonal line across the diagram represents the rate of rise of temperature of an inert body during the constant input of heat. Line (B) illustrates passive negative feedback such as might occur on a watery planet as a result of increasing cloudiness. Line (A) illustrates passive positive feedback such as could take place when an ice covered planet reached the melting point of water and its albedo changed from near 1.0 to a much lower value associated with open oceans and crustal rock. Line (C) is for an active feedback system with the goal of maintaining a set temperature.
With the passive negative feedback some constancy is achieved but at a value arbitrarily set by the properties of water and which cannot be changed. With the active system, constancy is possible at any chosen value. Positive feedback is used constructively so as rapidly to reach the chosen level and negative feedback used to keep the constancy. Furthermore the chosen set point can itself evolve as part of a more intricate system of evolutionary change.
Waler B. Cannon coined the term “homeostasis” for those coordinated physiological processes which maintain most of the steady states of a living organism. Homeostasis is very much an active process; one in which any departure from the chosen state is sensed and the difference between preference and reality amplified and used to oppose the perturbation and so restore the status quo. The stability of such a system, the quality of its homeostasis, is measured by its capacity to withstand perturbations. Now let us see how active regulation of a planetary scale might be achieved by the biota without the need for them to have foresight or receive divine assistance.
Daisy world
The dominant plant life of daisy world are black and white daisies. They are grazed by grey cattle but both producers and consumers flourish when the climate permits. Both species of daisy are identical in every respect other than the colour of their flowers and their growth varies with temperature in the same way. Because they absorb more radiation the local temperature of a stand of black daisies will always be higher than that of a stand of white daisies. As a result the rates of growth of the two species will be different at any given intensity of sunlight. To model this planetary ecosystem let us assume that the two species of daisy have a growth rate (β) which varies with temperature parabolically as follows:
β = AT – BT2
Where (A) and (B) are constants chosen so that growth is zero at below 5 and above 35 degrees Celsius and a maximum at 20 degrees. These limits are those that determine the growth of most contemporary vegetation. Under cool conditions the growth of the black daisies, which are locally warmer than are the white, will be favoured. Under hot conditions the white daisies will have the advantage. The rate of spread of one species into the zone of the other is given by a relationship described and experimentally confirmed. Carter and Prince (2), as follows:
dy/dt = β xy – γ y
Where (x) is the number of susceptible sites for growth and (y) is the number of infective sites. β and γ are the growth and death rates respectively.
Daisy world is a cloudless planet with no greenhouse gases. Figure 4 illustrates the response of the mean planetary temperature with increasing solar luminosity. The dotted line illustrates the temperature of a barren planet and the solid line when the daisies are present. The nodel illustrates how the powerful capacity of life to grow until a niche is full acts as an amplifier and natural selection a sensor in a control system which is able effectively and precisely to regulate planetary temperature at close to the optimum for the specified life form. No foresight or planning is required by the daisies only their opportunistic local growth when conditions favour them.
This type of model is not limited to the very artifical conditions of daisy world. A more general version would take into account the possibility that once life appears on a planet geochemical evolution will be limited by the circumscribed set of physical and chemical constraints which characterize the biota. Any external or internal chemical or physical change away from this set of conditions will lead not only to adaptation but also to the selection of those organisms whose growth alters the environment in so as to oppose the unfavourable development.
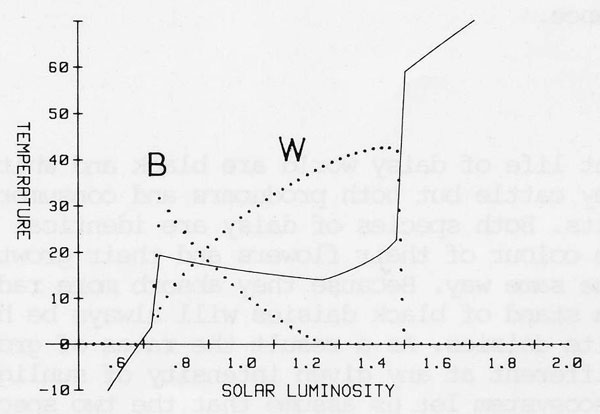
Figure 4. The active regulation of planetary temperature during the increase of solar luminosity by the growth and natural selection of two species of daisy. White daisies (W) albedo 0.7 black daisies (B) albedo 0.3 and the albedo of the bare planetary surface taken as 0.5. The solid line is the mean planetary temperature. The vertical axis also denotes the proportion of planetary area covered by each specie.
Past, present and future
The Gaia hypothesis postulates the existence of active systems for the chemostasis and thermostasis of the planetary environment. It predicts that the environment has been and will be stable and constant in spite of perturbations, whether sudden or from some continuous and progressive change. Strong support for the hypothesis will therefore come from the discovery of instances where there was clear evidence of the rapid and effective restoration of homeostasis after known perturbations.
There are several examples of major perturbations during the course of the history of life on Earth. The first and possibly the greatest was the origin of life itself. Whatever the environment was before life it should have changed profoundly soon after the planet was colonised. We know that conditions were favourable at the start of life so it is worth asking the question: “how was it that these conditions remained favourable in spite of the inevitable changes in surface, ocean and atmospheric composition imposed by life?” Thus the use of CO2 as a carbon source by early photosynthetic life could have gravely disturbed the precarious radiation balance of the Earth when warmed by a cooler sun.
It is interesting therefore to speculate on the composition of the Archean atmosphere after life began. There is evidence to suggest that the rate of carbon burial in the Archean was not greatly different from now. Much of the carbon entering present day anoxic sediments is returned to the atmosphere as methane. If the Archean production of methane was comparable with that which is now produced and bearing in mind that none of it would be oxidised by consumers in the anoxic Archean oceans, the flux to the atmosphere could have been substantial. In addition to methane, sulphur gases and possibly even nitrous oxide may also have been flowing into the Archean atmosphere.
It would need a general circulation model to predict such an atmosphere in detail. But it can be speculated that the troposphere would have a net reducing tendency with abundant polyatomic organic gases present. The radiative properties of such an atmosphere would solve the infra red problem of less CO2 and also serve if necessary to filter out ultra violet radiation at wavelengths less than 300 nanometres. The well established glaciation coincident with the first appearance of oxygen in the atmosphere at 2.2 giga years ago is consistent with the oxidative destruction of these organic gases.
The emergence of oxygen as an atmospheric gas may have been the largest perturbation the Earth has yet experienced. Significantly it was internally and biologically driven. The process of photosynthesis whereby oxygen and carbon are segregated and some of the carbon becomes buried, ineluctably drove the planetary surface ever more oxidising during the Archean. It is true that seme of this tendency was offset by the return of reducing materials by tectonic processes but until the critical problems posed by the presence of gaseous oxygen began to exert their effect on selection oxidation proceeded unchecked.
Among the minor but startling problems posed by the appearance of oxygen would have been the speciation of the element uranium. In the reduced form uranium is safely locked as water insoluble material dispersed a great dilution. In an oxidising environment uranium is water soluble and readily concentrated by microorganisms. This task was once successfully completed by microorganisms resident at a region which is now Gabon in Africa about 2.2 giga years ago. As a result a nuclear reactor commenced operation and ran for several million years. At that time uranium was substantially richer in the fissionable isotope U235 than now. It is fortunate that the Earth did not become oxidising in the early Archean for then the uranium was enriched close to bomb quality. Spectacular nuclear fireworks might have been more than the infant Gaia could have withstood.
By the time the metazoan biota were well established the presence of charcoal in the sediments provides a fossil record of ancient fires. The range of atmospheric oxygen over which fires can take place yet not be so devastating as to threaten all standing vegetation is 15% to 25% by volume. It is therefore tolerably certain that atmospheric oxygen has never ranged beyond these bounds in the last several hundred millions of years. This is a truly remarkable feat of regulation for in the previous 90% of the Earth’s history the pE has risen by at least ten units but is now held precisely constant. The mechanism by which oxygen is regulated is not yet known although it has been proposed by Watson et al. (17) that the control of the proportion of carbon buried in the anoxic sediments and hence the oxygen abundance is achieved through the regulation of the venting of methane to the atmosphere. Interestingly fires themselves exert a positive feedback on oxygen since carbon as charcoal is resistant to digestion by microorganisms and hence more is buried.
Throughout the existence of life on Earth there have been frequent collisions with planetisimals several kilometres in diameter, the most recent, 65 Myrs ago. The impact energy of these collisions is vast enough to have caused major, albeit temporary environmental changes, and was proposed as the cause of species extinctions in the fossil record, Alvarez et al. (1) . Figure 5 illustrates the impact craters so far discovered on the Canadian shield. Most of the events recorded by the craters represent an energy yield at the Earth’s surface about 108 times larger than the detonation of the present global stocks of nuclear weapons. Although the consequences of these impacts are not yet known in detail they do act as impulse tests of the “black box” system. If and when a detailed description of the sequence of events at one of these collisions is uncovered it will chart the course of the perturbation and the rapidity and effectiveness of the return to an optimum environment which follows. It could provide important evidence about the existence and the nature of Gaia.
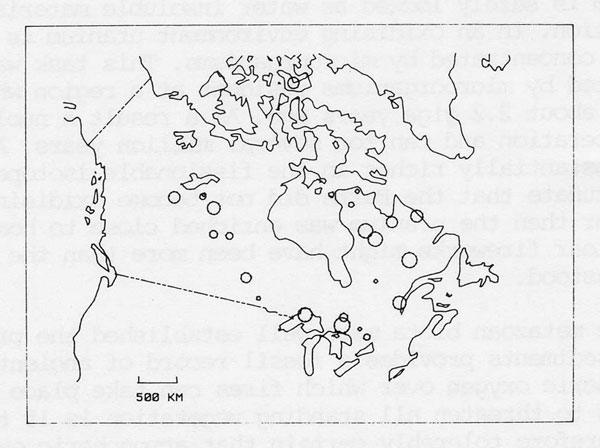
Figure 5. Map showing the impact craters so far discovered on the Canadian Shield.
A progressive change in the environment which spans the past, present and the future is that which relates to the climate of the Earth. One of the more certain conclusions of astrophysics is that stars increase their radiation flux as they age. There is a consenus among astronomers that the sun was very probably about 25 less luminous at the Earth’s origin than it is now. We know from the geological record that fluid water has always been present and from the origin of life that the climate cannot have been very different from now 3.5 giga years ago. In this context glaciations represent only minor departures from climatic constancy. Walker et al. (16) proposed that a progressive decrease in atmospheric CO2 from about 10% abundance at the start of life to the present 0.03% could through a decreasing greenhouse effect compensate for the progressive increase of solar luminosty. Although by itself carbon dioxide does not provide a very effective greenhouse, on the Earth its influence is amplified by presence of abundant water vapour.
The mechanism by which CO2 is varied inversely with the solar luminosity so as to maintain a constant temperature is the weathering of exposed calcium silicate rock. This is the only major sink for CO2 from the atmosphere and the rate of weathering has a positive temperature coefficient. Walker’s proposal provides a plausible abiological mechanism for climatic and CO2 regulation although with the present information on the fluxes of CO2 it appears to be only partially able to account for the constancy of the climate throughout the Earth’s history.
I do not disagree with the general basis of this interesting abiological control mechanism which would make Gaia redundant but wondered instead how much better it would work if life was included as a part of it. The real world is not abiological and the weathering of calcium silicate is very much a biological concern. At all levels from prokaryotic microorganisms to large trees and soil moving animals the biota participates in the process of rock digestion. The partial pressure of CO2 in the soil is 10 to 40 times greater than it is in the air. CO2 is actively pumped from the air by the biota to those regions of the soil where it can react with calcium silicate particles. The rate of CO2 fixation by plants is a strong function of both temperature and light intensity.
The need to have the biota participate in such a system is best illustrated by considering the consequences of its absence. If all life were deleted the soil CO2concentration would rapidly fall to below the present atmospheric level and weathering would be substantially reduced in rate. The input of CO2 to the atmosphere from volcanoes is on average constant and consequently the atmospheric CO2 concentration would rise until the current rate of weathering was re-established. The new equilibrium level would probably be above the current levels of the soil, about 1%. This is because diffusion from the air is very slow compared with the active penetration of the soil by plant roots. The ambient temperature under these conditions was calculated by Lovelock and Watson (10) to be about 20 degrees Celsius higher than now. The higher temperature might increase the abiological weathering rate but only slightly if it is limited by the rate of diffusion of CO2 to the calcium silicate rock.
Furthermore as recently modelled by Shukia and Minz (14) the lack of land life would so disturb the planetary water vapour transport that large areas would become desert where the weathering rate would be much reduced. It is also significant that CO2 and carbohydrate species transport to and from the oceans is very dependent upon the presence of life. The deletion of life from the oceans would lead to a further increase in atmospheric CO2.
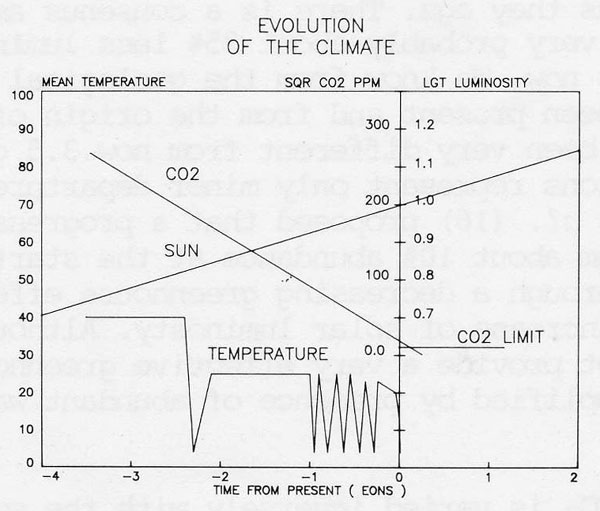
Figure 6. Evolution of the climate showing the variation of solar luminosity from its present value taken as 1.0. Also illustrated on the same time scales are the proposed decline in carbon monoxide concentration, expressed as the square root of its concentration in ppmv and the approximate range of mean surface temperatures in degrees Celsius.
Atmospheric CO2 abundance and climate is a current environmental concern as a result of the geochemically minor perturbation attributable to fossil fuel combustion. Figure 6 from Lovelock and Whitfield (11) summarizes in simple diagrammatic form the rise of solar luminosity during the Earth’s history and the fall of CO2 abundance needed to compensate for this rise in heat flux.
The most interesting feature of this diagram is its suggestion that a new perturbation of major magnitude is, on a geological ttmescale imminent. If the climate is to stay constant at near the optimum for the biota then the CO2 must be further reduced. However, a reduction below 100 ppm could not be suffered by most of the contenporary photosynthesisers. The diagram illustrates that such a level will be reached in only 100 Myrs. If zero CO2 were tolerable even this is approached in 200 Myrs.
It is unlikely that our descendants will be there to witness this interesting period when it comes. The past history of the Earth suggests that a near optimal planetary environment will be sustained by some other means.
References
1. Alvarez, I.W., Alvarez, W., Asaro, F., and Michell, H.V.: 1980, Science 208, p.1095.
2. Carter, R.N., and Prince, S.D.: 1981, Nature 293, p.644.
3. Dawkins, R.: 1982, The Extended Phenotype, Freeman, New York, p.114.
4. Ford Doolittle, W.: 1981, Coevolution 29, p.58.
5. Gilbert
6. Hitchcock, D.R., and Lovelock, J.E.: 1966, Icarus 7, p.149.
7. Holland, H.D.: 1978, The Chemistry of the Atmosphere and Oceans, Wiley Interscience, Chichester, U.K.
8. Hutchinson, G.E.: 1954, Biochemistry of the Terrestrial Atmosphere, in the Solar System (ed. Kuiper), Chapter 8, University of Chicago Press.
9. Lovelock, J.E., and Margulis, L.: 1974, Tellus 25, p.1.
10. Lovelock, J.E., and Watson, A.J.: 1982, J. Planet. Sci., in press.
11. Lovelock, J.E., and Whitfield, M.: 1982, Nature 296, p.561.
12. Margulis, L., and Lovelock, J.E.: 1974, Icarus 21, p.471.
13. Redfield, A.C.: 1958, Am. Sci. 46, p. 205.
14. Shukia, J., and Minz, Y.: 1982, Science 215, p.1498.
15. Sillen, L.G.: 1966, Tellus 18, p.198.
16. Walker, J.C.J., Hays, P.B., and Kasting, J.F., J. Geophys. Res., in press.
17. Watson, A.J., Lovelock, J.E., and Margulis, L.: 1978, Biosystems 10, p.293.