
By J. E. Lovelock, Department of Cybernetics, University of Reading, Reading, UK, and A. J. Watson, Marine Biological Association, Citadel Hill, Plymouth, U.K.
Published in Planet. Space Sci., Vol. 30, No. 8, pp. 795-802, 1982. (Received 12 September 1981)
Paper presented at the IAMAP / ICPAE Symposium “Origin and Evolution of Planetary Atmospheres”, 17-18 August 1981, Hamburg, West Germany.
Abstract: This is a review of the Gaia hypothesis which postulates a condition of planetary homeostasis affecting chemical composition and climate. Some criticisms are answered and a new model is introduced for the long term regulation of the mean surface temperature through the biological control of CO2 partial pressure.
Introduction
Theories of the climate and the chemical cornposition of Earth’s surface either ignore the presence of the biosphere or assign to it a passive role; a sort of non-participating spectator at a demonstration of physics and chemistry.
Redfield (1958), Rubey (1951), Hutchinson (1950), Berkner and Marshall (1966) and Sillen (1966) all in their time proposed a more active biological presence but their views, though respected, do not seem to have influenced mainstream science. The climate and the chemical composition of the Earth are still modelled as if their regulation were independent of the presence of the biota. It is as if the steady state motion of a bicycle and its rider were modelled with the rider a mere dead weight. It is true that the term “biogeochemical cycle” has recently become fashionable usage, but even here the rider is still merely something through which raw materials and energy flows, steering is not permitted.
During the 1960s the search for life on other planets, particularly Mars, led to a new approach to planetary biology. Lovelock (1965) and Hitchcock and Lovelock (1966) proposed that the presence of chemically based life on a planet would so change the composition of its atmosphere away from the abiological steady state that the change would be recognisable even at astronomical distances. The infra-red astronomy of P. and J. Connes (1966) found Mars to have an atmosphere dominated by CO2 and not greatly departed from the expectations of equilibrium chemistry. According to the new theory such a planet would not be expected to bear life. By contrast the Earth’s atmosphere was so far departed from the chemical equilibrium state as to advertise the presence of life even if viewed from as far away as the edge of the solar system. The coexistence of the two gases oxygen and methane at their present steady state concentrations was by itself near certain evidence for the presence of life. When the whole ensemble of atmospheric gases was viewed in this way it seemed that the interaction between life and the atmosphere is so intense that the atmosphere could be regarded as an extension of the biosphere. Not living but a construction chemostatically maintained by the biota. This thought led to the Gaia hypothesis. Lovelock and Margulis (1974), Margulis and Lovelock (1974) which postulates that the climate and the chemical composition of the Earth’s surface are kept in homeostasis at an optimum by and for the biosphere.
This paper briefly reviews the Gaia hypothesis and then develops from it a model mechanism for the long term regulation of atmospheric carbon dioxide and climate.
The Gaia hypothesis
This hypothesis and some of the evidence for it and criticisms of it, can be briefly stated as follows.
The presence on a planet of a global cybernetic system with the capacity to regulate the chemical composition of its atmosphere can be recognised from the extent to which a planet with such a system differs from the expected abiological steady state. This approach arises from Boltzman’s expression for entropy (S) in terms of the probability (P) of a molecular distribution:
S = k In P.
The very low entropy associated with living systems suggests a model configuration of extreme improbability compared with the expected and predictable steady state abiological background. If highly improbable molecular distributions extend beyond the boundaries of living entities so as to include also their planetary environment then the environment and life taken together can be considered to constitute a single larger entity. It is this operating system of life and its environment which we are calling Gaia. Effects due to the presence of life are not limited to chemical change. If life determines the abundance of a polyatomic gas such as CO2 then the planetary radiation balance will be altered. An effect such as this could be part of a system of planetary thermostasis.
Partial pressures
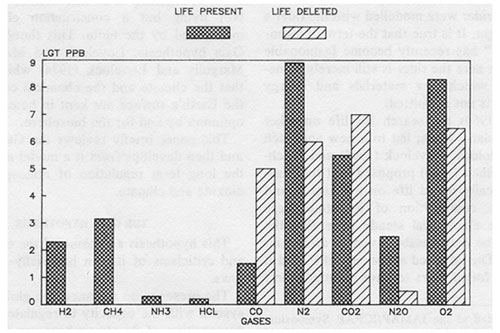
Figure 1: The abundance of gases in the present atmosphere compared with that expected for the abiological steady state.
In Fig. 1 the abundance of atmospheric gases are compared for the present Earth and for an Earth from which life had been deleted. The deletion is assumed to have taken place without any concomitant change in chemical composition or climate and these properties were then allowed to proceed to the abiological steady state. In Fig. 2 a similar comparison is made for the fluxes of the gases.
These two diagrams illustrate how much the present atmosphere of the Earth is in a transient and highly improbable steady state. Without life, gases such as methane and nitrous oxide would ; vanish in a few years and the major atmospheric gases nitrogen, oxygen and carbon dioxide would i reach a new and profoundly different equilibrium – in a million years or so. The evidence by which the presence of life on Earth is recognised is the simultaneous presence of gases such as oxygen and methane or of an easily decomposed gas such as nitrous oxide. There are no abiological processes capable of sustaining the fluxes or the abun’ dances of these disparate gases. The evidence from which the presence of Gaia is suggested is the maintenance of these reactive gases at a steady state composition.
Whitfield (1981) has commented on a similar disequilibrium in the elemental composition of the solutes dissolved in the oceans. Fig. 3 taken from Whitfield’s paper compares the biological fluxes of elements in the sea with the fluxes of the same elements from the inorganic environment. If the ocean was regulated according to the expectations of the abiological steady state all of the elements shown would be distributed in the lower left hand region of the diagram. As it is the reverse is found. Both toxic and nutritious elements are in disequilibrium.
Fluxes
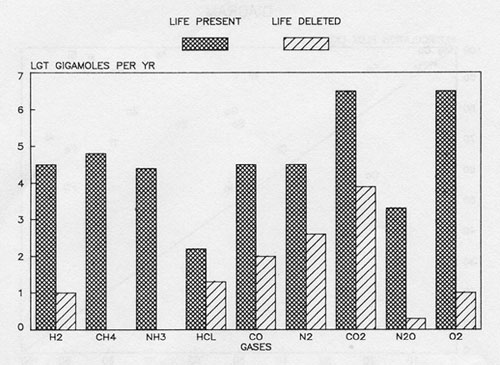
Figure 2: The fluxes of gases (giga moles per year) through the present atmosphere compared with those expected for the abiological steady state.
Kaplan (1975) objected to the use of chemical disequilibrium as evidence of the presence or influence of life and cited as examples the carbonaceous chondrites and the rocks of the lunar surface. In both of these rocks oxidised and reduced minerals coexist yet it is tolerably certain that they have never been touched by life. From this it was argued that even intense chemical disequilibria are equivocal as evidence for the presence of life. Kaplan’s criticism is relevant only for certain disequilibria in the solid state where events can be frozen, even for eons, witness the fossil bacteria of the archean, Tyler and Barghoorn (1954). In contrast, the disequilibria which are used as evidence for Gaia are of the liquid and gaseous states. They are observed to persist for periods much longer than the reaction times of the components considered. For example, the atmospheric concentrations of methane and of nitrous oxide have been observed for nearly 30 years to remain near to the steady state. There is no reason to believe that their abundance was substantially different from now at any time in the past ten thousand years, yet their atmospheric residence times are about 10 and 100 years, respectively.
The Gaia hypothesis has also been criticised on the grounds that Darwinian natural selection could never have led to the evolution of a single quasi immortal entity [Ford Dolittle (1981)]. This criticism is important for it concerns the form of Gaia. The critic envisages a new form of life, but we are postulating no more than a large control system which includes the biosphere as a component. Conventional neo-Darwinism sees evolution as the natural tendency of those species which leave the most progeny in an environment to be its inheritors. We do not disagree but remark that conventional wisdom also assumes the environment to be constant or if changed, only by external forces. What if there are environmental changes as a consequence of the evolution of the species themselves? If there is such an interaction then we have the recipe for a cybernetic system in which the species and the elemental cycles can evolve together perhaps seeking an optimum. There is no doubt that major changes in the Earth’s environment have been brought about by life itself. When the cyanobacteria finally completed their titration of the reducing equivalent of the Earth’s environment by separating and segregating the oxygen and carbon of CO2 this was a great evolutionary step. The planetary biology and the chemistry were simultaneously profoundly changed. If the links between life and its environment were closed, in a cybernetic sense, at this time, then the persistence of life through those hazards caused by the emergence of oxygen as an atmospheric gas is less puzzling.
Ocean flux diagram
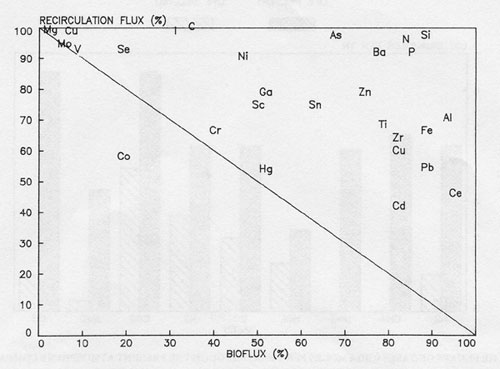
Figure 3: flux diagram showing the relationship between the percentage of the total removal of elements from the mixed zone attributable to the biota and the percentage total input provided by recirculation from the deep ocean.
If the distribution was abiologically controlled all points would lie along or beneath the diagonal.
One of the more convincing items of evidence in favour of Gaian regulation is the constancy of the climate since life began. During this period of 3.5 Ga it is thought that the sun has grown in size by 25% and there have certainly been substantial changes in atmospheric composition. What follows are some new thoughts on climate based on the assumption that the abundance of atmospheric CO2 plays an important part in its regulation and that the CO2 itself may be subject to Gaian manipulation.
The regulation of climate and CO2
Although astronomers are notoriously fickle with their speculations, the hypothesis that main sequence stars increase their luminosity as they age seems well founded. The evidence on which it is based is reviewed by Newkirk (1980) where also its applicability to models of the evolution of the Earth’s climate is discussed. As Gough (1980) comments, “It is an apparently straightforward consequence of basic atomic and nuclear physics, gas kinetics and Newtonian gravitation. Therefore the astrophysical conclusion is robust.”
Sagan and Mullen (1972) first commented on the paradox of a smaller sun coexisting with a climate equable enough for the origins of life. The sun was possibly 25 less luminous then than now, yet the geological record indicates the formation of sedimentary rocks as long ago as 3.8 Ga before the present. The persistence of life itself suggests that at no time did either freezing or near boiling conditions prevail. Contrary to the expectations of simple physics it is even suggested, Knauth and Epstein (1976) that the Archean climate was considerably warmer, with a surface temperature between 325 and 345 K.
It is usual to explain the early warmth by postulating the presence of some molecular species able to absorb and return to the surface infra-red radiation. Sagan and Mullen proposed ammonia as the candidate “greenhouse” gas which could supplement the otherwise insufficient infra-red absorbtion of water vapour. Owen et al. (1979) preferred carbon dioxide at a partial pressure up to 1000 times the present level. Carbon dioxide at such concentrations is plausible for a young and tectonically active planet and it is photochemically much more stable than is ammonia. It is possible even that some at least of the early warmth could have come from the presence of persistent aerosols of polymeric material which would be more stable in a reducing or neutral atmosphere. Whatever the explanation of the warmth of those very remote times we are still faced with the problem of explaining the constancy, or even the decline, of the climate in spite of the probable increase of solar output. Walker et al. (1981) have proposed an interesting purely abiological feedback process whereby changes in the Earth’s temperature are resisted during the increase in solar flux by an automatic adjustment of the CO2 partial pressure. Briefly, an increase in temperature directly affects the rate of reaction between CO2 and calcium silicate rocks and may also increase rainfall. Acting together these two effects hasten weathering and promote a new and lower equilibrium pressure of CO2. Lower CO2 pressure reduces the rise in temperature which otherwise would result from increased solar luminosity. The input of CO2 from tectonic sources is assumed to be constant in the long term.
We shall never know whether or not this otherwise plausible mechanism would work on a lifeless Earth. But there is no doubt that the contemporary rate of weathering of silicate rocks is biologically not geochemically determined. The partial pressure of CO2 in the soil where weathering occurs is 10 – 40 times higher (Holland, 1978) than is the atmospheric level. These high partial pressures of CO2 are maintained by soil bacteria; furthermore the rate of CO2 production by soil bacteria doubles for each 10 degree rise in temperature. It is probable that weathering has been influenced by the soil bacteria throughout many geological periods possibly as far back as the archean. Perhaps during the archean when solar luminosity was low, CO2 high, and the environment anoxic the effect of the soil bacteria was to reduce the soil CO2 and so sustain longer a high atmospheric CO2.
A test for Gaia is to consider what would happen if life were now deleted from the Earth. At first the partial pressure of CO2 in the soil would fall to a level below that of the atmosphere, since diffusion would limit the rate of its transfer to the silicate particles of the soil, which are the sink. In consequence the rate of weathering would be that set by a soil CO2 > 30 times less than now. Atmospheric CO2 would then rise in abundance until a new equilibrium was reached at which weathering proceeded at near to the present rate. It would have required at least 1% CO2 in the soil and rather more in the atmosphere since there would be a concentration gradient from the air to the silicate particles of the soil. It is assumed that, in the long term, the rate of input of CO2 from volcanic sources remained constant.
In order to investigate the effects on surface temperature of such a change we need a radiation model. We chose to use the quasi-grey, “equivalent radiative atmosphere” formulation due to Chamberlain (1980). This is a simplified approach and we have not included possible feedbacks due to ice or cloud cover. The transmission of the atmosphere is approximated by a step function and we use the power law dependencies given by Pollack (1969) to calculate the opacity due to CO2 and H2O using 13 wavelength intervals in the far infra-red. The method tends to underestimate and gives a temperature rise of 1.5°C in surface temperature for a doubling of CO2, at constant relative humidity. This compares with the 2°C change predicted from more detailed calculations. We compared the predictions of this simple method with the calculations of Owen et al. (1979) for the past evolution of surface temperature, with increasing solar flux and a decreasing CO2 partial pressure. For atmospheric CO2 pressures of less than 0.03 bar the two models agreed sufficiently well to justify the use of the simple calculations for a first order estimate. Table 1 illustrates the temperature increment to be expected if the present day levels of CO2 were changed. With constant relative humidity, a concentration of 1 gives an increment of over 20°C, an increase certain to have profound implications for the contemporary biota.
Table 1: Effect of CO2 changes

From a Gaian viewpoint these arguments suggest that the current temperature is actively kept at a level well below that expected for the abiological steady state. Carbon dioxide is pumped out of the air by the biota which use the accelerated weathering processes of the soil bacteria. This process will be assisted by the mechanical and chemical break down of rocks by plant roots, fungi and lichens and by the numerous invertebrates of the soil It is a very Gaian mechanism in which the geochemical feedback process proposed by Walker et al. (1981) is amplified and then used opportunistically by the biota.
Although highly specialised microorganisms can survive in hot springs at temperatures up to 90°C, the critical upper limit of temperature for main stream life is 56°C. If we accept the possibility that the climate is biologically controlled then the current low temperatures and the repeated glaciations since 2.3 Ga ago point towards the present temperatures of the Earth as near optimal for the current biosphere. The high internal temperatures of the homeotherms, 35 – 40°C, is not inconsistent with this since these animals are well adapted to much lower ambient temperatures.
Evolution of the climate
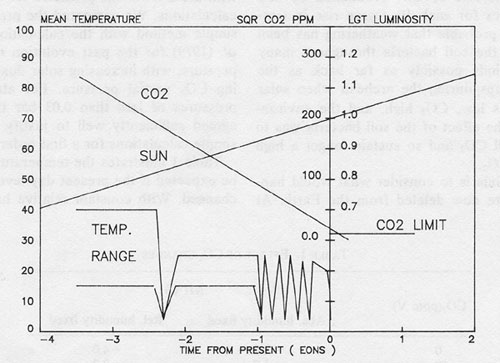
Figure 4: the variation of solar luminosity, relative to its present value taken as 1.0, with time expressed in giga years before and after the present time.
Also illustrated on the same time scale the approximate mean surface temperature in °C and the square root of the CO2 concentration (-ppm V).
Figure 4, from Lovelock and Whitfield (1981), illustrates the decline of CO2 abundance, the growth of solar luminosity and the mean surface temperature of the Earth from the archean to the present. Extrapolations of these properties into the near future are also illustrated. The solar luminosity changes are from Newkirk (1980), the CO2 partial pressure during the archean was taken to be that proposed by Owen et al. (1979). The mean surface temperatures in the past are not accurately known and the line has qualitative significance only. Glaciations define more certain temperatures and the notches in the line are at the times of known glaciations. We assumed as correct the proposals by Walker et al. (1981) and by Owen et al. (1979), that the principal means by which the Earth has kept a constant warmth, as the sun increased luminosity, was by reducing CO2. Some form of monotonic relationship is then needed to link the 7-10% abundance of archean CO2 with the present 0.03%. Our simple model suggested that for CO2 abundances above the present level, a constant mean temperature could be kept if the square root of the CO2 abundance was a function of the solar luminosity. Walker suggests a slightly different relationship that of the 0.364 power instead of the square root but both intersect the limiting CO2 level of 150 ppm close to 100 Ma from now.
If CO2 control is now the major process of climate regulation and if the Sun is indeed growing larger, then it seems that there is little further room for manoeuver. Table 1 shows the temperature decrement which would result from lowering the CO2 concentration to 50 ppm, well below the level which is presently needed by the biota to sustain rapid photosynthesis. Even complete removal of the atmospheric CO2 lowers the temperature by only a few degrees. In the future, it is probable that mean surface temperature will be forced to increase in step with solar output.
Future climates
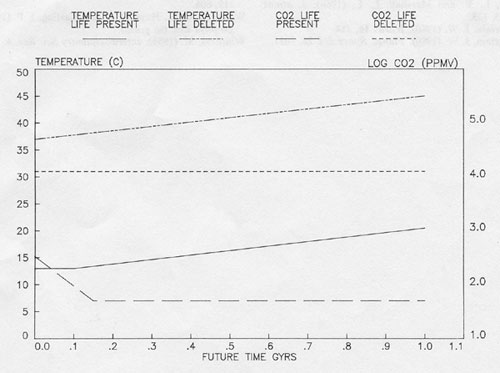
Fig. 5. the variation of mean surface temperature with solar luminosity for fixed relative and fixed absolute humidities.
The CO2 is held constant at 50 ppm V.
Figure 5 is a speculative look at the future evolution of the climate and CO2. It illustrates the trend of the mean surface temperature, calculated using our simple radiation model. Two bounds are shown, one for the continued operation of climate control down to 50 ppm CO2 at constant relative humidity and the maintenance of this CO2 level thereafter. The other bound assumes that Gaian regulation fails and the geochemical control takes over. Should this happen the CO2 would rise to between 1% and 2% and the temperature by 25°C. Such a world does not seem fit for the present biosphere. The relatively recent evolution of the C4 plants which can nourish at the 10 to 100 ppm level of CO2 suggest that adaptation to the lower CO2 levels of the lower bound may be possible and that the end is still distant for contemporary life when viewed from a human time scale. But when compared with the age of the biosphere, more than 3.5 eons, the time remaining seems short and if control fails is probably to be measured in hundreds rather than thousands of millions of years.
Mitchell (1976) expressed the occurrence of climatic episodes during the Earth’s history as a power series. Prominent among the recurring events is one with a time constant near 200 Myrs. This has been attributed by McCrea (1981) to the various possible climatic consequences of the passage of the solar system through the spiral arms of the galaxy, prominent among these consequences are the impact of plantesimals, such as the one described by Alvarez et al. (1980). Dilke and Gough (1972) offer an alternative or additional explanation, namely the inherent overstability of the Sun’s core which leads to episodes of mixing and a change of luminosity at intervals of a few hundred Ma. Whichever or both of these phenomena may occur, they have the capacity to induce heating as well as cooling of the Earth’s surface. Their importance could increase as the control of climate by CO2 abundance moved close to its lower limit.
The astronomers prediction of a growth of solar output is not a certainty. The Sun may have had a constant or only slightly varying luminosity. If this is so there is still the puzzling question of the anomalously low partial pressure of CO2 in the atmosphere. Is the steep concentration gradient of CO2 between the air and the soil or even the interior of an animal an accident of geochemistry; or is it still part of a Gaian system for climate regulation to cope with the lesser fluctuations imposed by minor internal and external changes.
References
Alvarez L. W., Alvarez, W., Asaro, F. and Michell, H. V. (1980). Science, N.Y. 208, 1095.
Berkner, L. V. and Marshall, L. L. (1966). J. atmos.Sci. 23, 133.
Chamberlain, J. W. (1980). Icarus 10, 314.
Chamberlain, J. W. (1980). Planet. Space Sci. 28, 1011.
Connes, J. and Connes, P. (1966). J. Opt. Soc. Am. 56,896.
Dilke, F. W. W. and Gough, D. O. (1972). Nature, Lond. 240, 262.
Ford Doolittle, W. (1981). Coevolution Q. 29, 58.
Gough, D. (1980). Nature, Lond. 288, 639.
Hitchcock, D. R. and Lovelock, J. E. (1966). Icarus 7, 149.
Holland, H. D. (1978). The Chemistry of the Atmosphere and Oceans, Wiley Interscience, Chichester, U.K.
Hutchinson, G. E. (1954). “Biochemistry of the terrestrial atmosphere”, in The Solar System (Ed. Kuiper), Chapter 8, University of Chicago Press.
Kaplan, I. R. (1975). Proc. R. Soc. Lond. B189, 180.
Knauth, L. P. and Epstein, S. (1976). Geochim. cosmochim. Acta 40, 1059.
Lovelock, J. E. (1965). Nature, Lond. 207, 568.
Lovelock, J. E. and Margulis, L. (1974). Tellus 26, 1.
Lovelock, J. E. and Whitfield, M. (1981). Nature, Lond. (in press).
McCrea, W. H. (1981). Proc. R. Soc. Lond. A375, 1.
Mitchell, J. M. (1976). Q. Res. 6, 481.
Newkirk, G. (1980). Proc. Conf. on Ancient Sun, (Ed. Pepin, R. O., Eddy, J. A. and Merrill, R. B.)
Owen, T., Cess, R. D. and Ramanathan, V. (1979). Nature, Lond. 277, 640.
Pollack, J. B. (1969). Icarus 10, 314.
Redfield, A. C. (1958). Am. Sci. 46, 205.
Rubey, W. W. (1955). Bull. Am. Geol. Soc. 62, 1111.
Sagan, C. and Mullen, G. (1972). Science. N. Y. 177, 52.
Sillen, L. G. (1966). Tellus 18, 198.
Tyler, S. A. and Barghoorn, E. S. (1954). Science, N.Y. 119, 606.
Walker, J. C. G., Hays, P. B. and Kasting, J. F. (1981). J. Geophys Res. (in press).
Whitfield, M. (1981). Interdisciplinary Sci. Rev. 6, 12.